Reward-space · Stability · Receptor Geometry · Ancient Neural Evolution
The mind does not move through a fixed inner landscape. It moves through a field that is continuously being shaped — by opportunity, by stability, and by the ancient neuromodulators that tune both.
Mind-state space may be imagined as a terrain of attractor basins, transition paths, thresholds, gradients of value, exploratory corridors, and regions of temporary stability.
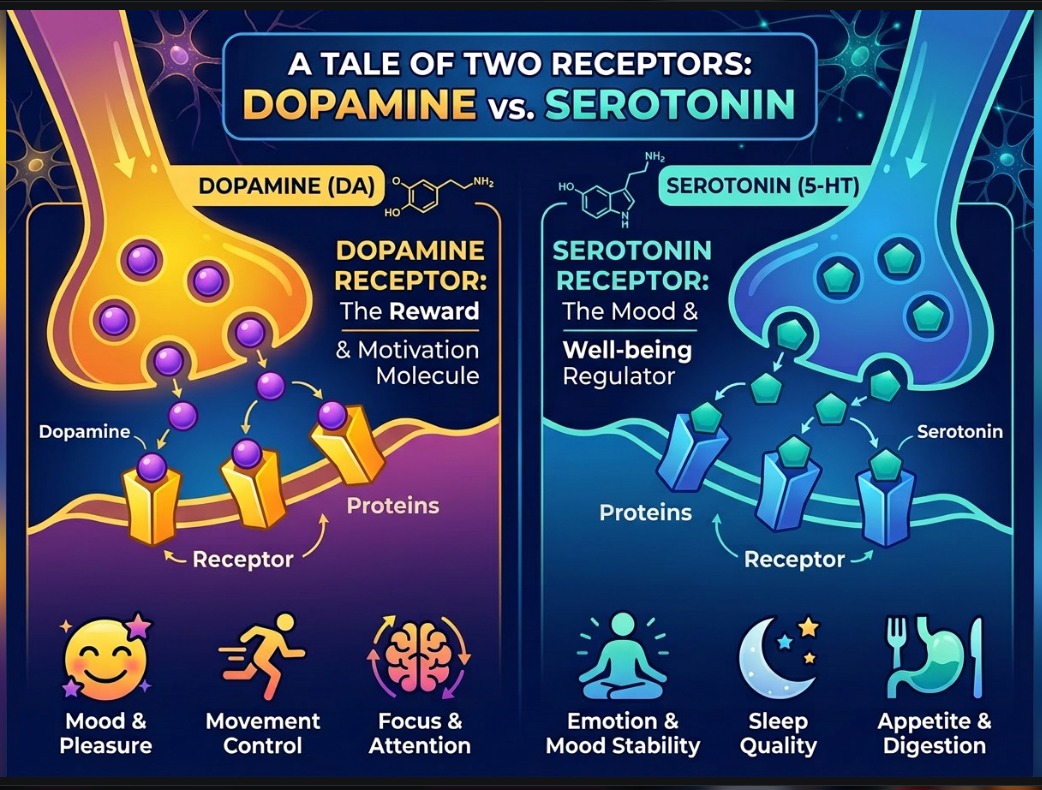
Dopamine and serotonin are often described as chemicals of reward, pleasure, mood, or balance. But they do something deeper. They do not merely carry signals through the nervous system; they help shape the field within which thought, feeling, motivation, and choice unfold.
attractor basins or stable states
pathways of transition
barriers and thresholds
gradients of value
zones of exploration
regions of temporary stability
In this view, consciousness is not only awareness within the landscape. It is also the felt movement through a landscape that is being tuned as one lives.
Evolutionary backdrop with the small inset retained inside the sea field
Dopamine is more than a reward chemical. It is a chemistry of movement, approach, exploration, and pursuit. It can deepen certain attractor basins, make value gradients steeper, lower transition barriers, and widen the field of exploration.
Dopamine edits possibility.
In Wolfram Schultz’s framework, reward is broader than pleasure. It includes whatever the organism treats as valuable for learning and behavior — food, water, relief, predictive cues, money, tokens, or status signals. Over time, dopamine shifts from the reward itself to the cue that predicts reward. The mind learns not only to enjoy, but to anticipate.
Serotonin, by contrast, edits persistence. It helps stabilize states, smooth abrupt shifts, and extend the horizon of value. It reduces impulsive jumps, strengthens continuity, and supports the capacity to remain rather than merely react.
Serotonin edits stability.
If dopamine says “Move,” serotonin says “Hold.”
Dopamine
deepen · open · energize · expand
Serotonin
stabilize · smooth · constrain · sustain
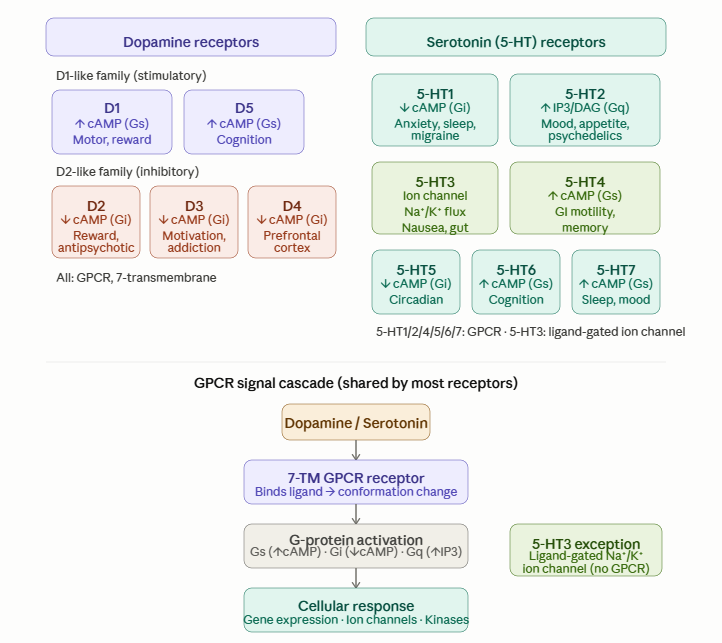
Dopamine and serotonin receptor families with the shared GPCR signal cascade
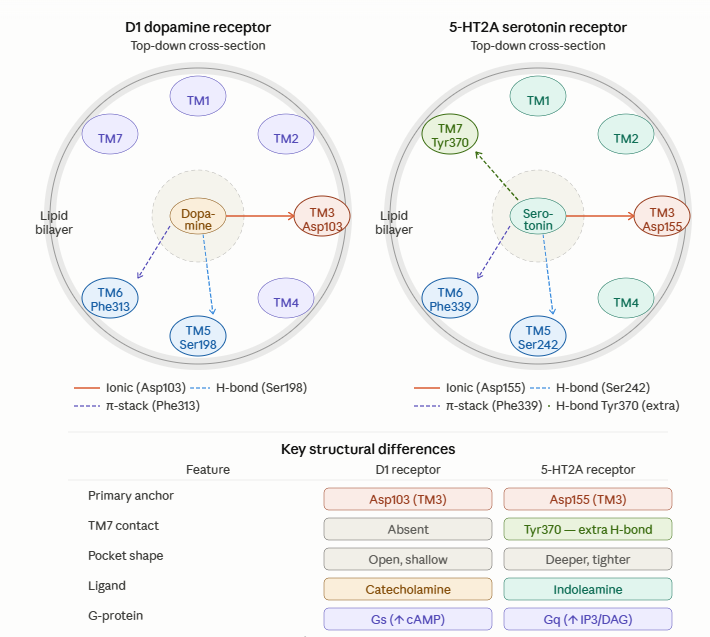
This complementarity appears even at the molecular scale. Dopamine is a catecholamine; serotonin is an indoleamine. Both bind to monoamine receptor systems with shared core features, yet serotonin receptors such as 5-HT2A form a deeper and tighter binding pocket than dopamine receptors such as D1.
Dopamine receptors divide broadly into D1-like receptors, generally stimulatory, and D2-like receptors, generally inhibitory. Serotonin receptors are even more diverse, spanning multiple signaling families, with 5-HT3 standing apart as an ion channel.
This diversity helps explain why dopamine and serotonin influence reward, motivation, inhibition, mood, cognition, and altered states in such different ways.
D1 dopamine receptor versus 5-HT2A serotonin receptor
IV
Molecular Context
Neurotransmitter Families · Molecular Form · Chemical Neighbourhood
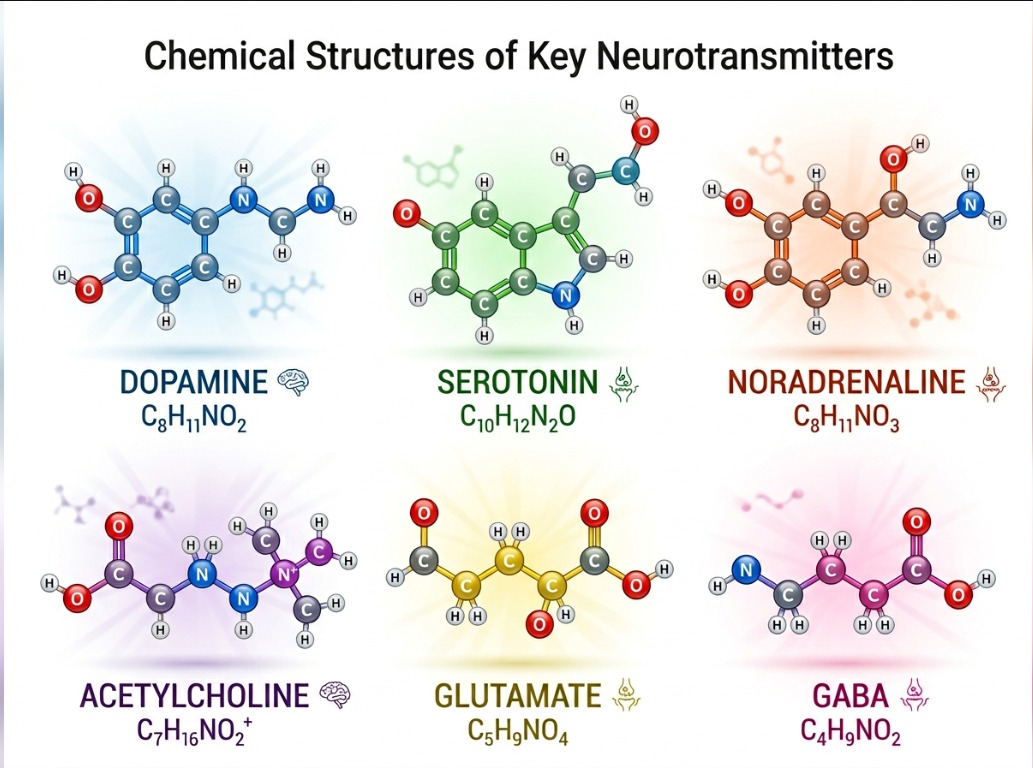
Seen in the wider molecular family, dopamine and serotonin sit among other neurochemical modulators that shape arousal, inhibition, learning, stress, and circadian balance. Their significance lies not only in isolated function, but in the network of biochemical relations that make conscious regulation possible.
chemical structure of key Neurotransmitters
V
Self as a Maintained Region
Occupancy · Coherence · Revision · Preservation
The self may not be a single attractor. It may be a maintained region within a neuromodulated field. Identity may be the persistence of a stabilized pattern: what is repeatedly entered, what is protected from disruption, what is reinforced across time, and what remains coherent amid change.
Dopamine contributes to updating. Serotonin contributes to holding. Together they mediate the moving balance between pursuit, preservation, and revision.
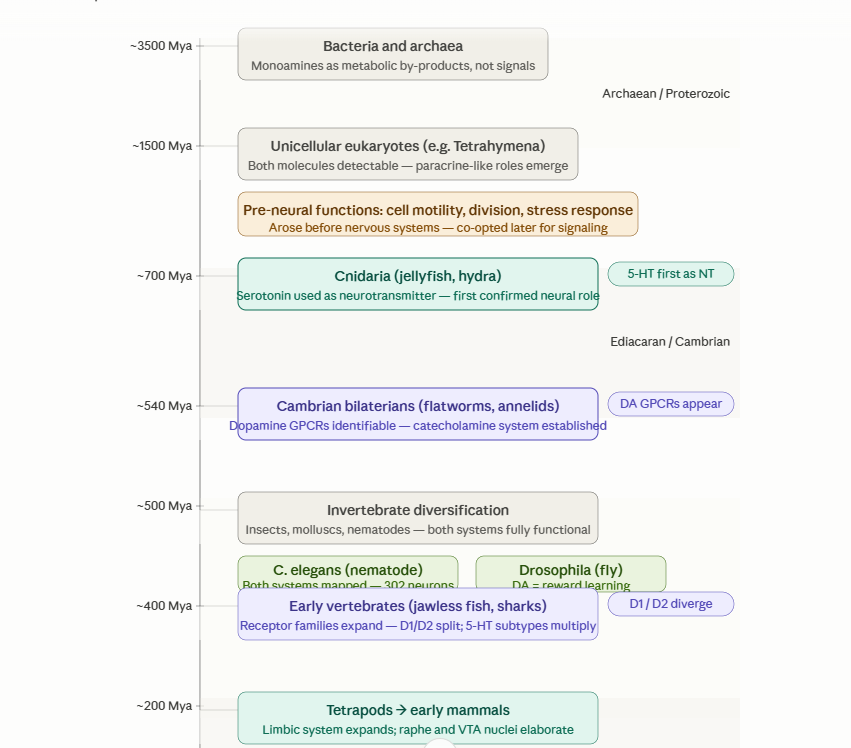
The story of dopamine and serotonin is far older than the human brain. Their roots go back deep into biological history, long before complex nervous systems emerged. Once neurons appeared, these molecules became tools for coordinating action and gradually entered the architecture of evolving nerve-net circuits.
Early stages — ancient chemistry before the modern brain
In early organisms such as sponges and cnidarians, organization was largely circular and radial. There was no true directional vector to manage, no forward axis in the later bilaterian sense. These forms could sense and respond, but not yet move through the world in a strongly directed way.
A major evolutionary threshold arose when the apical system and the blastoporal system came into association at the cephalic end. From that conjunction emerged the earliest groundwork of hypothalamic order — a first landmark stone on the great highway of neural evolution. Only then did organized exploration and exploitation of the environment become possible.
In this broad frame, dopamine may be seen as the neuromodulator of outward movement — of approach, seeking, pursuit, and engagement.
The blastoporal system, by contrast, used serotonin in relation to the movement-control of the coelomic domain, the future digestive system. It signaled satiety once energy requirement had been achieved. In that sense, it laid an early foundation for rest and digest — the precursor of the parasympathetic drives later expressed through vagal networks.
It is therefore not surprising that we now see a renewed fascination with parasympathetic regulation — vagal tone, affect modulation, biofeedback, and the monitoring of HRV through wristbands and rings. What appears today as a modern therapeutic fashion may actually be the latest expression of a very ancient biological logic.
The evolutionary story here is one of the most striking in all of neuroscience: both dopamine and serotonin are far older than brains.
Before neurons (3500–700 Mya). Both molecules appear in bacteria — not as signals, but as accidental metabolic by-products of amino acid chemistry. Tyrosine and tryptophan are ancient amino acids, and the enzymatic steps that convert them into catecholamines and indoleamines are chemically straightforward. Single-celled eukaryotes like Tetrahymena produce measurable amounts of both, using them for rudimentary purposes: regulating cell division, motility, and stress responses. This means serotonin and dopamine existed for roughly a billion years before any nervous system evolved to exploit them.
The co-option into neural signaling (700–540 Mya). The critical transition happened in early animals. Cnidarians (jellyfish, hydra) — which have nerve nets but no brain — are the first organisms where serotonin is confirmed as a genuine neurotransmitter, driving coordinated behavior like swimming and feeding. Dopamine's clear neural role appears slightly later, in bilateral animals of the Cambrian, where recognizable GPCR sequences for catecholamines emerge. The brain didn't invent these molecules — it inherited them and rewired existing chemistry into communication.
Later stages — vertebrate elaboration, primates, and the modern brain
With vertebrates came receptor expansion, specialized neuromodulatory hubs, and richer neural integration. By the time primates emerged, these systems had become deeply involved in cognition, planning, social behavior, and abstract thought.
What began as chemistry became regulation. What became regulation entered motivation, cognition, and selfhood.
VII
When the Geometry is Distorted
Depression · Addiction · Mania · Compulsion
If dopamine and serotonin help shape the landscape of mind, then many disorders may be understood as distortions within that landscape.
depression: flattened reward gradients and over-deep negative basins
addiction: hyper-deep local wells and compulsive return loops
mania: over-opened transitions and weakened constraints
compulsion: sticky cycles with reduced flexibility
In this view, pathology is not only chemical imbalance. It is distortion in the attractor ecology through which the mind moves.
Between them emerges a living geometry of reward, persistence, transition, and self-maintenance. Mind may be understood not merely as activity in the brain, but as movement through a neuromodulated landscape of attractors.
And consciousness, in part, may be the felt life of that landscape.